Cell and Molecular Biology and Imaging of Stem Cells features original and review articles written by experts who have made significant contributions to stem cell biology and imaging. Chapters cover a broad spectrum of aspects of the field, including Stem cells from the Amniotic Fluid and Placenta, Biomaterials as Artificial Niches for Pluripotent Stem Cell Engineering, Low-Intensity Ultrasound in Stem Cells and Tissue Engineering, Mammalian Neo-Oogenesis from Ovarian Stem Cells, Oct4-EGFP Transgenic Pigs as a New Tool for Visualization of Pluripotent and Reprogrammed Cells, Regulation of Adult Intestinal Stem Cells, Stem Cell Therapy for Veterinary Orthopedic Lesions, Sex Steroid Combinations in Regenerative Medicine for Brain and Heart Diseases, Hair Follicle Stem Cells, The Potential of Using Induced Pluripotent Stem Cells in Skin Diseases, Mitochondrial Differentiation in Early Embryo Cells and Pluripotent Stem Cells, and The Role of Centrosomes in Cancer Stem Cell Functions.
Cell and Molecular Biology and Imaging of Stem Cells addresses a wide variety of cell and molecular topics in unprecedented detail, and is a must-read for graduate students and academic and industry professionals in the expanding field of stem cell biology.
Reviews new imaging tools and markers for cell and molecular imaging in stem cell biology.
Covers novel aspects of stem cell imaging in reproductive biology and stem cell niches
Includes chapters on the developing area of centrosome biology as it applies to embryonic and adult stem differentiation
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Chapter 1 Cell and Molecular Biology and Imaging of Stem Cells: Stem Cells from the Amniotic Fluid and Placenta
Amritha Kidiyoor, Sean V. Murphy, and Anthony Atala
Wake Forest Institute for Regenerative Medicine, Wake Forest University School of Medicine, North Carolina, USA
Introduction
The promise of stem cell technology has mainly focused on the potential of pluripotent or multipotent cells to differentiate into a variety of different cell types representative of all three germ layers (Dushnik-Levinson and Benvenisty, 1995). Embryonic stem cells (ESCs) were the first truly pluripotent stem cell type described and subsequent research has provided a wealth of knowledge relating to stem cell development and lineage commitment (Reubinoff et al., 2000; Brivanlou et al., 2003). The main disadvantages of using ESCs for cell therapy is the potential for immune rejection of the allogeneic source of cells and their potential to form tumors in vivo. In the past few years, researchers have described methods to induce somatic cells into pluripotent stem cells by a process known as reprogramming (Takahashi and Yamanaka, 2006; Takahashi et al., 2007; Wernig et al., 2007; Stadtfeld et al., 2008; Zhou et al., 2009). This involves introducing transcription factors, proteins or small molecules into terminally differentiated cells to induce reprogramming into a stem cell phenotype. These induced pluripotent stem cells (iPSCs) possess many of the same properties as ESCs in terms of self-renewal and pluripotency. The advantage of using iPSCs is that patient-specific cell lines can be generated for autologous cell therapies. However, like ESCs, iPSCs possess the potential to form tumors in vivo, so are likely to be most useful for in vitro applications.
Adult stem cells are stem cell populations that have been identified in organs and tissues of adults such as the bone marrow (Ballas et al., 2002), blood (Lewis and Trobaugh, 1964) brain (Taupin, 2006), lung (Giangreco et al., 2002), and heart (Kinder et al., 2001). Adult stem cells are tissue-specific progenitors and are mainly involved in repair of their corresponding organ. They possess the ability to self-renew like ESCs and are more appealing to use as a replacement therapy, as they circumvent ethical concerns. In addition, autologous transplantation is possible with adult stem cells. The main disadvantage is that adult stem cells often require invasive biopsies for cell isolation, and the resultant cell populations are generally restricted to generating cell lineages corresponding to their organ of origin (Weiner, 2008). An attractive alternative is stem cells that can be isolated from gestational tissue such as the placenta, amnion membrane, and the amniotic fluid (De Coppi et al., 2007; Troyer and Weiss, 2008; Serikov et al., 2009; Murphy et al., 2010; Galende et al., 2010). Gestational tissue is usually discarded after birth and usage of this tissue involves minimal ethical or legal concerns. Amniotic fluid is frequently obtained in the second trimester during amniocentesis to detect any chromosomal abnormalities, malformations, and also to determine the sex of the fetus (Joo, 2011). Gestational tissue such as the placenta, placental membranes, and amniotic fluid are untapped reserves of stem cells. In the past decade, research groups have isolated and characterized stem cell populations that are highly multipotent, with the ability to differentiate into hematopoietic, osteogenic, chondrogenic, adipogenic, endothelial, myogenic, neural, and lung cells, among other cell lineages (In ‘t Anker et al., 2003, Portmann-Lanz et al., 2006; De Coppi et al., 2007). These cells also possess potent immunomodulatory properties, such as production of anti-inflammatory factors as well as interacting with immune cells to modulate the immune response (Murphy et al., 2010, 2011). These properties make peri-natal stem cells an attractive alternative for cell therapy. Therefore, the use of peri-natal stem cells for regeneration or replacement of damaged or diseased tissue such as blood and immune system (Ottersbach and Dzierzak, 2005; Ditadi et al., 2009), bone defects (Fan et al., 2012), myocardial infarction (Zhao et al., 2005; Bollini et al., 2011), neural degeneration (Kakishita et al., 2003; De Coppi et al., 2007), lung disease (Carraro et al., 2008), and diabetes would be valuable (Wei et al., 2003; Chang et al, 2007).
Gestational Tissue Development
Fetal placental tissue is derived from the trophoblast layer of the blastocyst, and functions to provide nutrients, eliminate waste, and provide gas exchange via the mother’s blood supply. The placental membrane provides a protective sac for the embryo and its contents. The inner cell mass of the developing embryo forms the epiblast and hypoblast layers. The hypoblast gives rise to extraembryonic tissue, while the epiblast gives rise to the ectoderm, mesoderm and endoderm, germ cells and extraembryonic mesoderm of the yolk sac, amnion, and allantois (Gardner and Beddington, 1988; Loebel et al., 1988; Downs and Harmann, 1997; Downs et al., 2004). Cells of the epiblast collected before implantation are the source of ESCs and can generate the entire fetus (Benitah and Fyre, 2012). The allantois tissue forms the umbilical cord and the mesenchymal part of the mature placenta (Downs and Harmann, 1997; Moser et al., 2004).
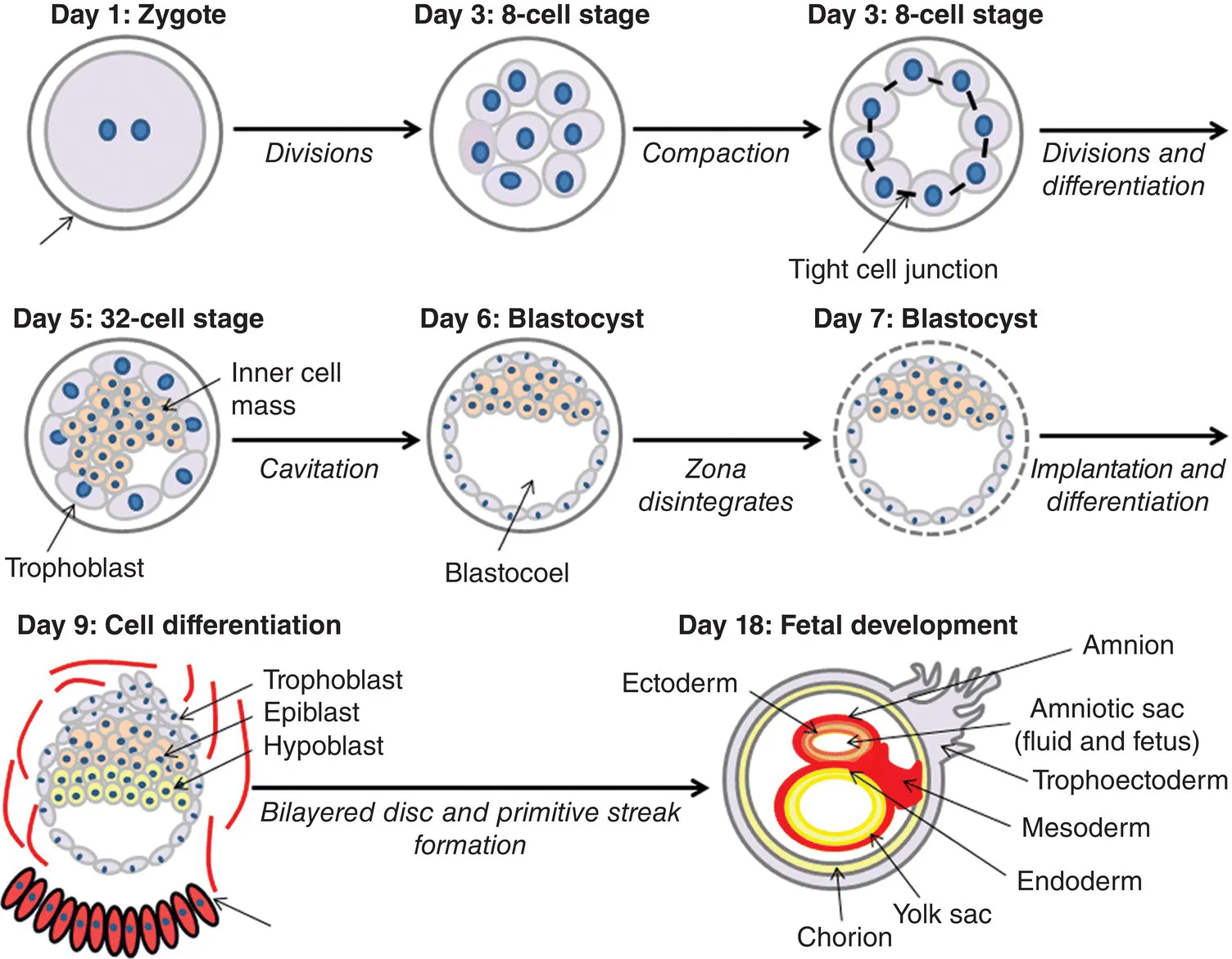
Around day 8.5 of gestation the hypoblast and epiblast form a bilayered disc and separate the blastocyst into two chambers; the yolk sac and the fluid-filled amniotic sac (Parameswaran and Tam, 1995; Kinder et al., 1999) (Figure 1.1). The amniotic sac consists of a pair of thin membranes: the outer membrane, the chorion, which envelops the inner membrane and forms a part of the placenta; the inner membrane, the amnion, which envelops the developing fetus and amniotic fluid (Kinder et al., 1999; Robinson et al., 2002).
Figure 1.1. The initial stages in the development of an embryo. A single-celled zygote undergoes multiple cell divisions, at the eight-celled stage the cells undergo compaction and bind tightly to one another. Further cell divisions and cell differentiation lead to the formation of the morula (32-celled stage), followed by cavitation resulting in the development of a blastocyst. The zona pellucida disintegrates, followed by implantation. The trophoblast layer of the blastocyst gives rise to the fetal placenta. Further divisions and differentiation occurs giving rise to the primitive streak and hence the three layers of the embryo. The epiblast and hypoblast form a bilayered disc that leads to the formation of amnion and chorion membranes.
The amniotic fluid is initially an isotonic fluid consisting of nutrients promoting fetal growth: carbohydrates, proteins, lipids, phospholipids, urea, and electrolytes. As the fetus develops, urine is excreted, which changes the composition and volume of the amniotic fluid (Heidari et al., 1996; Srivastava et al., 1996; Sakuragawa et al., 1999; Bartha et al., 2000). Amniotic fluid ensures movement of the fetus and symmetrical structural fetal development and growth. Amniotic fluid is inhaled and swallowed by the fetus contributing to lung development and gastrointestinal tract development respectively. Amniotic fluid contains a variety of cell types present during development, such as cells sloughed off the developing fetus, the alimentary tract, respiratory tract, urogenital tract, fetal amnion membrane, and skin (Medina-Gómez and del Valle, 1988).
The mature human placenta has both a fetal as well as a maternal component. Much of the placenta originates from the inner cell mass of the morula (chorion, amnion, and mesenchymal core of the chorion), with the trophoblast layer contributing towards the outer layer of the placenta. As the origin of gestational tissue or extraembryonic tissue takes place during the very early stages of embryonic development, these cells possess multipotent differentiation potential and hence are a rich and valuable source of stem cells for cell therapy.
Isolation and Characterization of Peri-Natal Stem Cells
Gestational tissue is usually discarded after birth so obtaining tissue is non-invasive and resourceful. Multipotent cells can be isolated from gestational tissue: placenta (Steigman and Fauza, 2007), amnion membrane (Alviano et al., 2007), chorion membrane (Bailo et al., 2004), and amniotic fluid (Kaviani et al., 2001) (Figure 1.2 ). Mesenchymal stromal cells (MSCs) can be isolated from the ...
Table of contents
Cover
Title page
Copyright page
Contributors
Preface
Chapter 1: Cell and Molecular Biology and Imaging of Stem Cells
Chapter 2: Biomaterials as Artificial Niches for Pluripotent Stem Cell Engineering
Chapter 3: Low-Intensity Ultrasound in Stem Cells and Tissue Engineering
Chapter 4: Mammalian Neo-Oogenesis from Ovarian Stem Cells In Vivo and In Vitroa
Chapter 5: Oct4-EGFP Transgenic Pigs as a New Tool for Visualization of Pluripotent and Reprogrammed Cells
Chapter 6: Regulation of Adult Intestinal Stem Cells through Thyroid Hormone-Induced Tissue Interactions during Amphibian Metamorphosis
Chapter 7: Stem Cell Therapy for Veterinary Orthopedic Lesions
Chapter 8: Sex Steroid Combinations in Regenerative Medicine for Brain and Heart Diseases
Chapter 9: Hair Follicle Stem Cells
Chapter 10: The Potential of Using Induced Pluripotent Stem Cells in Skin Diseases
Chapter 11: Mitochondrial Differentiation in Early Embryo Cells and Pluripotent Stem Cells
Chapter 12: The Role of Centrosomes in Cancer Stem Cell Functions
Index
End User License Agreement
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
Both plans are available with monthly, semester, or annual billing cycles.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go. Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Cell and Molecular Biology and Imaging of Stem Cells by Heide Schatten in PDF and/or ePUB format, as well as other popular books in Biological Sciences & Cell Biology. We have over 1.5 million books available in our catalogue for you to explore.