![]()
1
Introduction
The study of amphibians – both extinct and extant – makes a significant contribution to our understanding of how organisms develop and evolve. Like few other vertebrate groups, amphibians have been studied extensively from an early historic phase until today. Their modern exemplars have made an essential contribution to our understanding of phenomena such as morphogenesis, plasticity, larvae, metamorphosis, heterochrony, viviparity, feeding, ecology, speciation and microevolution, and – most recently and sadly – extinction. Their rich fossil record provides unique insights into ontogeny and paleoecology, phylogeny and macroevolution. Hence, the knowledge of amphibian evolution holds a pivotal position in the study of vertebrates.
Admittedly, amphibians are neither the most speciose, nor particularly spectacular vertebrates. They are often sluggish and slow, with a cold and moist skin covered with mucous and venom glands. Most of them are not very large, and many species are so tiny that they are easily overlooked. At the same time, amphibians are often the preferred objects for studies in development, ecology, and evolution. What, then, makes them such prominent study taxa? Why should their evolutionary history be of such wide general interest to biologists? There are historical reasons, influenced by their ready availability for study and the relatively easy breeding conditions of some laboratory taxa. However, amphibians are also special among vertebrates in many ways, not least in their capacity to survive and propagate in unstable environments, as well as in their ability to change from one habitat to a profoundly different one. Some amphibians have mastered the regeneration of organs in a way unthinkable in most other vertebrates, and they have repeatedly evolved live-bearing species, each time with different features. Some amphibians breathe with lungs, others with gills, and yet others through their skin – and many amphibians employ a combination of all these respiratory mechanisms. Finally, amphibians are a group whose evolutionary history dates back as far as the Early Carboniferous, a time span encompassing 330 million years of change and stasis, diversification and extinction, and fascinating examples of evolutionary innovation. It is the purpose of the present book to trace this history, seeking to understand features of amphibian evolution in the frameworks of development and ecology, the two major foci of modern evolutionary biology. It is the interdisciplinary questions that are the most fascinating in this field, and therefore the second major theme of the book is the question of how we conduct studies on the fossil record, development, ecology, and evolution of amphibians and beyond.
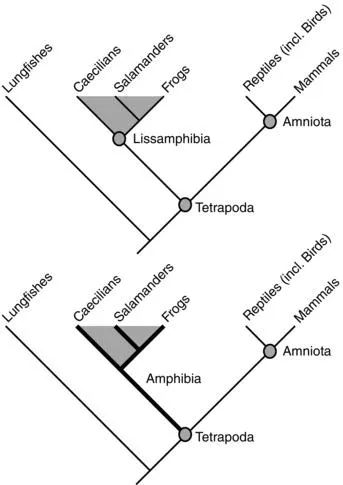
What is an amphibian? The phylogenetic definition that I will use is straightforward: any member of the three modern groups salamanders (Caudata), frogs (Anura), and caecilians (Gymnophiona) is an amphibian (Figure 1.1). The correct systematic name for that group is Lissamphibia, and all lissamphibians share a common ancestor that lived sometime in the Late Paleozoic (~330–290 myr).
There is a large gap between lissamphibians and the manifold Paleozoic and Mesozoic taxa commonly referred to as “amphibians.” Some of these must rank among the ancestors of lissamphibians, but authors still debate which taxa fall into the lissamphibian stem-group. To avoid confusion, it is reasonable to distinguish between the lissamphibian relatives (phylogenetically called “stem-amphibians”) and all other taxa. The others are referred to here as “early tetrapods” when their relationships to Lissamphibia and Amniota are uncertain, and as “stem-amniotes” if their affinity with amniotes can be made plausible. Here, I follow the majority view on the origin of Lissamphibia, which holds that temnospondyls, members of a speciose clade encompassing almost 300 species, form the stem-group of lissamphibians (Bolt 1969; Milner 1993; Ruta and Coates 2007; Sigurdsen and Green 2011; Maddin et al. 2012).
Therefore, when speaking of Paleozoic and Mesozoic amphibians, I refer to temnospondyls, and thus I employ a scheme in which Lissamphibia forms a subgroup within a larger clade Amphibia. The alternative views will be discussed in depth in Chapter 9 (phylogeny). Whereas this book deals mainly with lissamphibians and amphibians, it also tackles many problems concerned with early tetrapods.
1.1 Changing paradigms in amphibian evolution
Amphibians bear a most appropriate name in several respects, and the scientist who coined the term was probably not aware of all of them. Literally meaning “living on both sides,” the name points to the capacity to transform and adapt to divergent living conditions. In the narrow sense, the two sides are freshwater and land: the stereotyped amphibian life cycle includes the water-born newt or tadpole transforming into an adult land salamander or frog. Yet there are many other ways of amphibian existence, exemplified by the limbless caecilians, most of which live in the soil, the lungless and live-bearing salamanders, some of which ably climb trees, or the non-transforming axolotl, which is effectively a hypertrophied, sexually mature salamander larva. There are many more such cases, and on closer inspection one may even think there are as many different life cycles as there are species. These amazingly varied life histories differ far more than the slight variations in ontogeny known from other vertebrates. They often harbor built-in switches, responding to environmental inputs. Water conditions, temperature, food availability and properties, and oxygen form some of these factors, but there are many others, often confined to individual species or populations.
Amphibians are also peculiar because their fossil record is extraordinarily good. Although relatives of modern amphibians are often too small and delicate to be well preserved in most sediments, Paleozoic and early Mesozoic deposits yield a wealth of other, much larger amphibian fossils. These fossils tell us about a bizarre and alien world, playing in an exotic geographical setting and climate, and revealing highly unusual aspects of development and ecology. The abundance of early amphibians and their presence in numerous different deposits has made them preferred study objects for paleontologists ever since their first discovery in the 1820s. The most striking feature of these ancient forms is their huge size – ranging between 0.5 and 6 m. Compared with living amphibians, they had a very different morphology, many of them resembling modern crocodiles, while others reveal convergences to modern flatfishes, moray eels, giant salamanders, caecilians, and lizards.
In recent decades, discoveries of many new fossils have changed our view of early amphibians profoundly. Fossils are usually interpreted within the framework of phylogenetic hypotheses, spanned by well-known extant organisms. This procedure arrives at extant groups that give the best model for the understanding of the extinct group. In the case of amphibians and early tetrapods, the classic living model organisms were the modern salamanders, because of their apparently plesiomorphic appearance and the biphasic life cycle (larval–metamorphic). One might call this a central dogma in the study of tetrapod origins. Indeed, salamanders appeared to be perfect model organisms: their general body architecture, their “primitive” mode of locomotion on land, and the capacity of water-living larvae to transform into a terrestrial adult were seen as essential features of all early tetrapods. The central assumption was that the first tetrapods conquered land in the same way as many modern salamanders do it – namely, during metamorphosis.
Is the evolutionary conquest of land recapitulated in each baby salamander and frog? Formulations like that may be elegant, but have little to do with what really happened. There is no simple parallelism between ontogeny and phylogeny, let alone in such developmentally complex organisms as amphibians. The underlying processes are entirely different: stochastic selection on the evolutionary level, genetic and developmental mechanisms on the organism level. The whole issue of heterochrony, first triggered by such extraordinary cases as the axolotl, has become a multifaceted issue to analyze in recent years. New fossils, including those of Paleozoic baby amphibians, shed light on the life cycles of early amphibians (Boy 1974; Schoch 2009). These data amounted to the insight that metamorphosis was not shared by most of these early taxa, and that the salamander model is far from appropriate for the understanding of early tetrapods (Schoch 2002).
This model has also been challenged by many finds that indicate a more aquatic, fish-like habit of many early tetrapods (Coates and Clack 1990, 1991). These taxa (see Figure 1.2 for examples) retained lateral lines and gills as adults, and their skeletons were hardly capable of supporting longer excursions on land. The available evidence from fossil footprints confirms this, revealing that these animals were extremely slow when forced to cross dry land. They did not undergo a metamorphosis like modern amphibians. In many cases, adults are found in the same environments as their juveniles.
This touches the core of a second dogma on the fish–tetrapod transition, the ecological argument. The classic ecological scenario holds that tetrapods were attracted by food outside the water, that there must have been selection pressures driving their ancestors onto land. However, fossil evidence counters this idea by showing that early tetrapods and amphibians lived primarily in the water, retained many fish-like features and organs, and preyed on fish or other water-dwelling animals. New evidence from histology supports this conclusion, because many early tetrapods retained calcified cartilage inside their long bones to make their bodies heavier, while others had lightly built bones, providing excellent swimming but very poor walking abilities. In all of these taxa, the internal structure of limbs was not adapted to meet torsional stress such as that caused by locomotion on land (Sanchez et al. 2010). The old ideas of Alfred Sherwood Romer (1956, 1958), a pioneer in the study of early tetrapod evolution, are revived: then regarded as an oddity rather than mainstream opinion, his suggestion was that the origin of tetrapods took place under water, and that true land vertebrates appeared substantially later. Clearly, the salamander is not a reliable model for these long-extinct taxa. In turn, modern amphibians as a whole appear much more alien and interesting when these results are borne in mind. They form a separate, successive strategy to generate a land vertebrate, with many fascinating adaptations that were not features of early tetrapods, but evolved in the 330-million-year history of amphibian evolution after their split from the amniote ancestors. We are also more fully able now to trace some key aspects of this evolutionary pathway, although many problems are still unresolved.
The study of amphibian evolution – of extinct as well as extant taxa – reveals another very interesting aspect: ontogeny. In stark contrast to other groups of tetrapods, but similar to various fishes, amphibians are subject to profound ontogenetic change, reflecting a broad range of responses to environmental parameters. Although ancient taxa had very different ontogenies, they were sometimes as complex as modern ones. This reaches a stage at which it becomes necessary to consider the whole life cycle as a unit of taxonomy, phylogeny, ecology, and evolution. In paleontology, this concept has been put forward only recently. One outcome of these efforts is the present book, summarizing recent work and numerous still-unpublished observations. For paleontology, the life cycle concept means that single ontogenetic stages are not sufficient to trace evolutionary changes. Many problems in phylogenetic analyses result from the unsettled questions raised by ontogenies and developmental evolution. Fortunately, the preservation of different size classes in fossil amphibians provides insight into this field, permitting detailed comparisons between extant and fossil ontogenies. The old and troubled concept of heterochrony comes into mind almost automatically here: neoteny, in its classic example of the axolotl as a sexually mature larva. Yet the new field of developmental evolution (evo-devo) is much more than the study of ontogeny and phylogeny. As pioneered by Ivan Ivanovich Schmalhausen and Conrad Hal Waddington, it focuses on the phenotype as an active player, responding to environmental changes, resisting perturbation from inside and outside, and being able to remain remarkably stable throughout evolution if required. However, the more obvious capacities of amphibian phenotypes are their flexibility and plasticity. This covers the important aspect of the reaction norm, a concept uniting development and ecology under the evolutionary umbrella.
The significance of fossil amphibians for the understanding of evolution is obviously manifold: their own evolutionary history is full of detailed stories, their relationship to modern amphibians is complex and reveals many perplexing convergences, their paleoecology has many unique features and provides insight into habitats, environments, and climates long ago, and the connection between evolution and development has been studied extensively in some Paleozoic and Mesozoic clades. This leads to the recognition of metamorphosis, a key feature of modern amphibians, as a life history strategy that evolved some 300 million years ago. Finally, the bearing of early tetrapod fossils on the fish–tetrapod transition is profound and has the potential to further shift the picture.
1.2 Paleobiology: data, methods, and time scales
Although there is one true history of early land vertebrates that needs to be found, only aspects of this s...