Biological Sciences
Oxidative phosphorylation
Oxidative phosphorylation is the process in which energy is produced in cells through the transfer of electrons from electron donors to electron acceptors. This process occurs in the inner mitochondrial membrane and involves the generation of ATP, the cell's primary energy currency. It is a key component of cellular respiration and plays a crucial role in the production of energy for various cellular activities.
Written by Perlego with AI-assistance
Related key terms
1 of 5
10 Key excerpts on "Oxidative phosphorylation"
 eBook - PDF
eBook - PDF- Efraim Racker, Anthony San Pietro(Authors)
- 2014(Publication Date)
- Academic Press(Publisher)
PART II Formation of Adenosine Triphosphate in Particulate Systems This page intentionally left blank LECTURE 7 Oxidative phosphorylation IN MITOCHONDRIA Seek simplicity and then distrust it. —A. N. Whitehead Phosphorylation linked to electron transport processes in the Krebs cycle is commonly referred to as oxidative phos-phorylation. I shall follow this custom, but not without emphasizing once more that in substrate level phosphoryla-tion and photophosphorylation also, the energy of oxidative processes is used to generate ATP. STAGES OF DISCOVERY I should like to give you a brief outline of the history of discoveries in the area of Oxidative phosphorylation, which will serve as a gentle introduction to this complex problem. Four stages of development can be distinguished (Scheme 7.1), each of which lasted approximately 10 years. We are Stage 1. Recognition of the phenomenon Stage 2. Quantitative evaluations Stage 3. Localization of sites Stage 4. Resolution and reconstitution Scheme 7.1 History of the Study of Oxidative phosphorylation now in the fourth stage, and if we continue to follow this pattern in the future, the major problems in oxidative phos-phorylation should be solved by 1969. 83 84 7. Oxidative phosphorylation in Mitochondria The first stage—recognition of the existence of the phe-nomenon—started in 1930 with the studies of Engelhardt, 80 who recognized with remarkable insight the relationship between the processes of oxidation and phosphorylation. The next breakthrough came with the discovery by Kalckar, in 1937 and 1939, of respiration-dependent formation of ATP in cell-free preparations. 81 The second stage, that of quantative evaluations, began with the studies by Belitzer and Tsibakova 82 with minced muscle, and by Ochoa 83 with cell-free brain and heart prep-arations. These investigators observed that close to 2 moles of phosphate were esterfied per atom of oxygen consumed.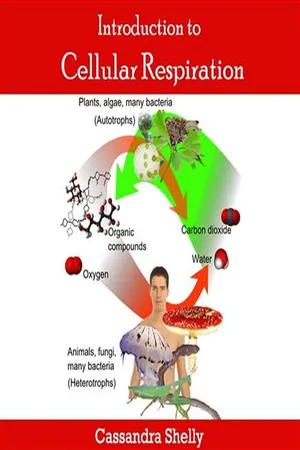 No longer available |Learn more
No longer available |Learn more- (Author)
- 2014(Publication Date)
- The English Press(Publisher)
________________________ WORLD TECHNOLOGIES ________________________ Chapter- 3 Oxidative phosphorylation The electron transport chain in the mitochondrion is the site of Oxidative phosphorylation in eukaryotes. The NADH and succinate generated in the citric acid cycle are oxidized, releasing energy to power the ATP synthase. ________________________ WORLD TECHNOLOGIES ________________________ Oxidative phosphorylation is a metabolic pathway that uses energy released by the oxidation of nutrients to produce adenosine triphosphate (ATP). Although the many forms of life on earth use a range of different nutrients, almost all carry out Oxidative phosphorylation to produce ATP, the molecule that supplies energy to metabolism. This pathway is probably so pervasive because it is a highly efficient way of releasing energy, compared to alternative fermentation processes such as anaerobic glycolysis. During Oxidative phosphorylation, electrons are transferred from electron donors to electron acceptors such as oxygen, in redox reactions. These redox reactions release energy, which is used to form ATP. In eukaryotes, these redox reactions are carried out by a series of protein complexes within mitochondria, whereas, in prokaryotes, these proteins are located in the cells' inner membranes. These linked sets of proteins are called electron transport chains. In eukaryotes, five main protein complexes are involved, whereas in prokaryotes many different enzymes are present, using a variety of electron donors and acceptors. The energy released by electrons flowing through this electron transport chain is used to transport protons across the inner mitochondrial membrane, in a process called chemiosmosis . This generates potential energy in the form of a pH gradient and an electrical potential across this membrane. This store of energy is tapped by allowing protons to flow back across the membrane and down this gradient, through a large enzyme called ATP synthase. No longer available |Learn more
No longer available |Learn more- (Author)
- 2014(Publication Date)
- College Publishing House(Publisher)
________________________ WORLD TECHNOLOGIES ________________________ Chapter 8 Oxidative phosphorylation The electron transport chain in the mitochondrion is the site of Oxidative phosphorylation in eukaryotes. The NADH and succinate generated in the citric acid cycle are oxidized, releasing energy to power the ATP synthase. Oxidative phosphorylation is a metabolic pathway that uses energy released by the oxidation of nutrients to produce adenosine triphosphate (ATP). Although the many ________________________ WORLD TECHNOLOGIES ________________________ forms of life on earth use a range of different nutrients, almost all carry out Oxidative phosphorylation to produce ATP, the molecule that supplies energy to metabolism. This pathway is probably so pervasive because it is a highly efficient way of releasing energy, compared to alternative fermentation processes such as anaerobic glycolysis. During Oxidative phosphorylation, electrons are transferred from electron donors to electron acceptors such as oxygen, in redox reactions. These redox reactions release energy, which is used to form ATP. In eukaryotes, these redox reactions are carried out by a series of protein complexes within mitochondria, whereas, in prokaryotes, these proteins are located in the cells' inner membranes. These linked sets of proteins are called electron transport chains. In eukaryotes, five main protein complexes are involved, whereas in prokaryotes many different enzymes are present, using a variety of electron donors and acceptors. The energy released by electrons flowing through this electron transport chain is used to transport protons across the inner mitochondrial membrane, in a process called chem.-iosmosis . This generates potential energy in the form of a pH gradient and an electrical potential across this membrane. This store of energy is tapped by allowing protons to flow back across the membrane and down this gradient, through a large enzyme called ATP synthase. No longer available |Learn more
No longer available |Learn more- (Author)
- 2014(Publication Date)
- The English Press(Publisher)
____________________ WORLD TECHNOLOGIES ____________________ Chapter- 6 Oxidative phosphorylation The electron transport chain in the mitochondrion is the site of Oxidative phosphorylation in eukaryotes. The NADH and succinate generated in the citric acid cycle are oxidized, releasing energy to power the ATP synthase. Oxidative phosphorylation is a metabolic pathway that uses energy released by the oxidation of nutrients to produce adenosine triphosphate (ATP). Although the many ____________________ WORLD TECHNOLOGIES ____________________ forms of life on earth use a range of different nutrients, almost all carry out Oxidative phosphorylation to produce ATP, the molecule that supplies energy to metabolism. This pathway is probably so pervasive because it is a highly efficient way of releasing energy, compared to alternative fermentation processes such as anaerobic glycolysis. During Oxidative phosphorylation, electrons are transferred from electron donors to electron acceptors such as oxygen, in redox reactions. These redox reactions release energy, which is used to form ATP. In eukaryotes, these redox reactions are carried out by a series of protein complexes within mitochondria, whereas, in prokaryotes, these proteins are located in the cells' inner membranes. These linked sets of proteins are called electron transport chains. In eukaryotes, five main protein complexes are involved, whereas in prokaryotes many different enzymes are present, using a variety of electron donors and acceptors. The energy released by electrons flowing through this electron transport chain is used to transport protons across the inner mitochondrial membrane, in a process called chemiosmosis . This generates potential energy in the form of a pH gradient and an electrical potential across this membrane. This store of energy is tapped by allowing protons to flow back across the membrane and down this gradient, through a large enzyme called ATP synthase. No longer available |Learn more
No longer available |Learn more- (Author)
- 2014(Publication Date)
- The English Press(Publisher)
____________________ WORLD TECHNOLOGIES ____________________ Chapter- 1 Oxidative phosphorylation The electron transport chain in the mitochondrion is the site of Oxidative phosphorylation in eukaryotes. The NADH and succinate generated in the citric acid cycle are oxidized, releasing energy to power the ATP synthase. Oxidative phosphorylation is a metabolic pathway that uses energy released by the oxidation of nutrients to produce adenosine triphosphate (ATP). Although the many ____________________ WORLD TECHNOLOGIES ____________________ forms of life on earth use a range of different nutrients, almost all carry out Oxidative phosphorylation to produce ATP, the molecule that supplies energy to metabolism. This pathway is probably so pervasive because it is a highly efficient way of releasing energy, compared to alternative fermentation processes such as anaerobic glycolysis. During Oxidative phosphorylation, electrons are transferred from electron donors to electron acceptors such as oxygen, in redox reactions. These redox reactions release energy, which is used to form ATP. In eukaryotes, these redox reactions are carried out by a series of protein complexes within mitochondria, whereas, in prokaryotes, these proteins are located in the cells' inner membranes. These linked sets of proteins are called electron transport chains. In eukaryotes, five main protein complexes are involved, whereas in prokaryotes many different enzymes are present, using a variety of electron donors and acceptors. The energy released by electrons flowing through this electron transport chain is used to transport protons across the inner mitochondrial membrane, in a process called chemiosmosis . This generates potential energy in the form of a pH gradient and an electrical potential across this membrane. This store of energy is tapped by allowing protons to flow back across the membrane and down this gradient, through a large enzyme called ATP synthase. No longer available |Learn more
No longer available |Learn more- (Author)
- 2014(Publication Date)
- College Publishing House(Publisher)
____________________ WORLD TECHNOLOGIES ____________________ Chapter 9 Energy Transformations Oxidative phosphorylation The electron transport chain in the mitochondrion is the site of Oxidative phosphorylation in eukaryotes. The NADH and succinate generated in the citric acid cycle are oxidized, releasing energy to power the ATP synthase. ____________________ WORLD TECHNOLOGIES ____________________ Oxidative phosphorylation is a metabolic pathway that uses energy released by the oxidation of nutrients to produce adenosine triphosphate (ATP). Although the many forms of life on earth use a range of different nutrients, almost all carry out Oxidative phosphorylation to produce ATP, the molecule that supplies energy to metabolism. This pathway is probably so pervasive because it is a highly efficient way of releasing energy, compared to alternative fermentation processes such as anaerobic glycolysis. During Oxidative phosphorylation, electrons are transferred from electron donors to electron acceptors such as oxygen, in redox reactions. These redox reactions release energy, which is used to form ATP. In eukaryotes, these redox reactions are carried out by a series of protein complexes within mitochondria, whereas, in prokaryotes, these proteins are located in the cells' inner membranes. These linked sets of proteins are called electron transport chains. In eukaryotes, five main protein complexes are involved, whereas in prokaryotes many different enzymes are present, using a variety of electron donors and acceptors. The energy released by electrons flowing through this electron transport chain is used to transport protons across the inner mitochondrial membrane, in a process called chemiosmosis . This generates potential energy in the form of a pH gradient and an electrical potential across this membrane. This store of energy is tapped by allowing protons to flow back across the membrane and down this gradient, through a large enzyme called ATP synthase. eBook - PDF
eBook - PDF- G. E. W. Wolstenholme, David W. FitzSimons, G. E. W. Wolstenholme, David W. FitzSimons(Authors)
- 2009(Publication Date)
- Wiley(Publisher)
Release of ATP from a hydrophobic to hydrophilic environment may consume most of the energy. This paper is meant to give a brief introduction to our current knowledge of the interconversion of respiratory energy into phosphate bond energy during Oxidative phosphorylation in mitochondria. I shall summarize some facts and 23 24 M. KLINGENBERG models for the mechanisms underlying this important energy transduction process. Such a contribution appeared to be desirable in this symposium as no other paper deals with this major synthetic source of ATP in aerobically living eukaryotic cells. I am aware that this treatment cannot be complete and apologize for any omissions or one-sided views presented here. Energy transformation linked to Oxidative phosphorylation in mitochondria has been subject to intensive discussion and controversies over the years. Sometimes, exaggerated ideas have been propounded without sufficient experimental basis. We must realize that the mitochondrial energy transfor- mations are linked to membranes and that, therefore, the level of understanding is part of a general lack of comprehension of thefunctional processes in membranes. In other words, the research on Oxidative phosphorylation does not lag behind research on other membrane-linked functions such as on membrane-bound enzymes or on membrane carriers. On the contrary, the amount of experimental data available on the mitochondrial membrane far exceeds that for any other membrane. THE SYSTEM OF Oxidative phosphorylation The components comprising the system of Oxidative phosphorylation are largely known now. They may be divided into (a) the electron transfer com- ponents including dehydrogenases and the coenzymes, and (b) the ATP synthase complex. Both parts of the system are more or less tightly associated with the inner mitochondrial membrane and, therefore, the isolation and definition of the various protein constituents encounter great difficulties. eBook - PDF
eBook - PDF- Alasdair Steven, Wolfgang Baumeister, Louise N. Johnson, Richard N. Perham(Authors)
- 2016(Publication Date)
- Garland Science(Publisher)
Importantly, this led to the buildup of oxygen in the atmosphere after the vast amounts of Fe 2+ in the early seas had been oxidized to the Fe 3+ form. This created conditions for the fur-ther evolution of organisms capable of profiting from the large amounts of energy released by oxidative metabolism of the carbonaceous material derived from CO 2 fixation. The molecular machinery of respiratory chain phosphorylation is described in Section 15.2, that of photosynthesis is covered in Section 15.3, and the ATP synthase, a molecular motor driven by chemiosmosis, is explained in Section 15.4. 15.2 BIOLOGICAL OXIDATION AND THE RESPIRATORY CHAIN Biological oxidation generally consists of the removal of electrons or hydrogen atoms from a substrate and their transfer to an acceptor (see Box 15.1). In aerobic organisms, the NADH and FADH 2 generated by the oxidative metabolism of, say, glucose (see Figure 15.1) or fatty acids (Chapter 9) are reoxidized with molecular oxygen as the ultimate electron acceptor. The reducing equivalents are passed to O 2 along an electron transport chain ( ETC ) com-posed of a series of large multi-subunit protein complexes. These complexes are vectorially embedded in a membrane: the cell membrane in bacteria, or the inner membrane of the mitochondria in eukaryotes. In the uncompartmentalized cells of bacteria, the enzymes of carbohydrate and fatty acid oxidation are all located in the cytosol. In contrast, the enzymes of glycolysis in eukaryotes are in the cytoplasm but the enzymes of the citric acid cycle (including the PDH complex) Biological Oxidation and the Respiratory Chain 666 Chapter 15: Bioenergetics Box 15.1 Cofactors of biological oxidation Biological oxidation normally requires the removal of two H atoms or of one or more electrons from a designated sub-strate: typically, say, the oxidation of an alcohol to a ketone (R 1 R 2 CHOH → R 1 R 2 C=O + 2H) or the oxidation of the Fe ion in a cytochrome (Fe 2+ → Fe 3+ + e – ). eBook - PDF
eBook - PDF- Sankhavaram R. Panini(Author)
- 2013(Publication Date)
- Thieme(Publisher)
140 III Cellular Respiration Mechanism of Oxidative phosphorylation The success of Oxidative phosphorylation depends on the respira-tory chain’s ability to accomplish three key goals: 1. Transfer electrons from NADH and the reduced form of flavin adenine dinucleotide (FADH 2 ) to O 2 2. Establish a proton gradient across the inner mitochondrial membrane 3. Synthesize ATP 1. Transfer of Electrons from NADH and FADH 2 to O 2 The components of the electron transport chain are arranged in a manner that allows electrons to flow from the molecules with the lowest E o ¢ (e.g., NADH = –0.32 V; FADH 2 = –0.22 V) to that with highest E o ¢ (O 2 = + 0.82 V). Additional characteristics of electron transfer reactions of the electron transport chain are as follows: — Electrons are transferred along the electron transport chain in various forms, such as electrons (e – ), hydride ion (:H – ), and hydrogen atoms (·H). — Electron transfers are facilitated by pairs of molecules that serve as oxidizing and reducing agents (redox pairs), such as NAD + /NADH, FAD/FADH 2 , ubiqinone/ubiquinol, and the flavin mononucleotides (FMN/FMNH 2 ). — Electron transfers are also facilitated by redox couples, which are the oxidized and reduced forms of an element, such as Fe 3+ /Fe 2+ and Cu 2+ /Cu + . The difference in redox potential between acceptor and do -nor molecules ( D E o ¢ = E o ¢ acceptor – E o ¢ donor ) is associated with a free energy change ( D G ° ¢ ). D G ° ¢ can be calculated using the following equation, where n is the number of electrons transferred, and F (Faraday constant) is 96.5 kJ/volt·mol (23.06 kcal/volt·mol). D G ° ¢ = – n F D E o ¢ As the foregoing equation implies, D G ° ¢ and D E o ¢ are directly related but are opposite in magnitude. Thus, the greater the D E o ¢ , the more negative is the D G ° ¢ . No longer available |Learn more
No longer available |Learn more- Gustavo Blanco, Antonio Blanco(Authors)
- 2017(Publication Date)
- Academic Press(Publisher)
2+ homeostasis. Thereby, mitochondrial defects may also affect those processes.Respiratory Control
Like other metabolic processes, Oxidative phosphorylation requires the presence of adequate concentrations of the necessary substrates in the medium, including ADP, Pi , O2 , and a metabolite that by oxidation can donate electrons to NAD or FAD. When the availability of any of these four factors decreases, the synthesis of ATP is limited. The level of ADP in the mitochondrial matrix is very important as a regulator of Oxidative phosphorylation. When there is no ADP, respiration stops. In the presence of oxygen, and an adequate supply of oxidation substrates, mitochondrial respiratory activity depends on the availability of ADP. This requires a close relationship between electron transport and phosphorylation and has been called respiratory control . It is clear that the synthesis of ATP depends on the continuous flow of electrons from a substrate of oxidation to O2 . Furthermore, in intact mitochondria, electron flow occurs only while ATP is synthesized. This is explained by the coupling between electron transport and proton pumping across the inner membrane. The proton-motive force is responsible for the synthesis of ATP. If external protons cannot return to the matrix through the F0 channel piece, not only the production of ATP in the F0 F1 complex will be blocked, but also the passage of electrons through the respiratory chain will stop. Proton accumulation in the outside of the membrane will increase the gradient, reducing, and eventually stopping, the transfer of any additional protons. This regulatory mechanism is intended to limit mitochondrial oxidation to adjust it to the physiological requirements of ATP of the cells, preventing wasteful consumption of substrates.The respiratory chain is highly responsive and reacts very quickly to any demand. In general, the system maintains high levels of ATP in cells. Stimulation of ATP consuming activity, such as muscle contraction, immediately increases the production of ADP. Increasing levels of ADP in the cell activate substrate oxidation, electron transport, proton pumping, flow of H+ via the F0 F1
Index pages curate the most relevant extracts from our library of academic textbooks. They’ve been created using an in-house natural language model (NLM), each adding context and meaning to key research topics.